Someday.
Many of my freshman have not had much in the way of biology, and some concepts that are basic to future courses in the major are vital. Among them are transformation, antibiotic resistance, and gene regulation.
In the laboratory portion of Biology 111, we use the famous pGLO plasmid, which contains a transcriptional fusion between the araBAD promoter and green fluorescence protein (GFP). Thus, when the araBAD promoter is "active," GFP is made. And GFP is a wonderful thing for students to observe, due to its lovely color and the fact is is clearly evident with inexpensive UV "flashlights."
The restriction map of pGLO is pretty straightforward.
 |
| From: http://classroom.sdmesa.edu/eschmid/Lab7-B1.jpg |

It's interesting to discuss this process with students. The concept that the bacterium that takes up the plasmid DNA by transformation will then express the genes carried by the plasmid can sometimes take a bit of getting used to for students. But it is a strong foundation to base other concepts upon. I often say that the role of an introductory course is to create a "tree" of connected concepts on which branches and leaves can be placed later, in future courses. So it is with transformation and plasmids and drug resistances.
What I adore showing students is what happens later. One of the problems with ampicillin resistance in the molecular biology lab is that it is usually mediated by the secreted enzyme beta-lactamase, which cleaves the beta-lactam ring of penicillin-like drugs such as ampicillin. This in turn means that the concentration of ampicillin surrounding the transformant colonies decreases. So far, so good.
One of the things that I emphasize to students is that only a small proportion of the bacterial cells can successfully take up and express the plasmid DNA (compared to the cells that do not). And more importantly, that many antibiotics actually don't kill bacteria...but simply keep them from growing. Thus, with time, a transformation plate using ampicillin resistance can look like this.

I really am proud of this photo; it looks lovely to me.
The glowing central colonies (the original transformants, containing pGLO) have degraded much of the ampicillin surrounding them, allowing nontransformed cells to eventually grow. These are the famous "satellite colonies" that bedevil many molecular biologists who leave their plates out for the weekend before picking transformants! I have them pick a big colony, streak it out, and show that it both glows on arabinose medium and is ampicillin resistant. What amazes them is that the satellite colonies (that do not glow) are sensitive to ampicillin when tested the same way!
This is a fabulous concept question for final exams! It really illustrates the central concepts involved, and involves critical thinking.
Most of the time, people who use pGLO ignore the fact that it is essentially a reporter gene construct: linking GFP synthesis to the activity of the araBAD promoter of the arabinose operon of E. coli.
So I have a couple of issues I like to show students, as illustrated from a lecture slide below.

D and L arabinose are enantiomers of one another, and this is a good way to illustrate that "mirror images" of the same chemical can have quite different effects. Molecular shape---recognition---is all about that. And it is highly relevant to organic chemistry and pharmacology, as enantiomers of certain drugs can act quite differently from one another.
This idea is central to my course. Heck, I even give it a fake name: stericity. Molecular shape confers functionality. Alter that shape, and the functionality is altered, too.
Finally, when discussing gene regulation and operons, the idea of multiple levels of control sometimes confuses students (perhaps not yours; I can only discuss my own experiences in the classroom and laboratory). So I try to get students to think about the arabinose operon (because of the connection to pGLO, actually).
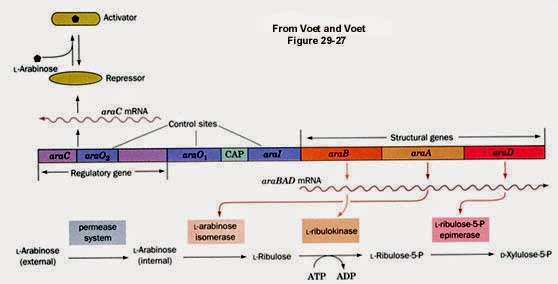 |
| From: https://www.bio.cmu.edu/courses/03441/AraGal/AraOverview.jpg |
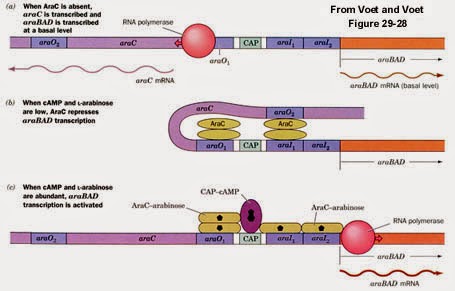 |
| From: https://www.bio.cmu.edu/courses/03441/AraGal/AraExpression.jpg |

And sure enough, on the combination plate (glucose + arabinose), GFP is turned off in the presence of glucose. Over the next week, guess what? GFP gets turn on and is visible on the combination plate. It's great to ask the students why (as the glucose is used up, cAMP levels rise, which form a complex with CRP, assisting in activation of the araBAD promoter).
I have also been known to put pGLO into E. coli deletion mutants in the crp and cya (adenylate cyclase) genes...to show the role of that system in gene regulation---putting it all together.
To summarize, it is very possible to use this humble plasmid to illustrate many central concepts in molecular biology and bacterial genetics. And because of the fluorescence of GFP, it is the kind of thing that students enjoy, and remember.
Now, if only I had similar constructs between araBAD and RFP or YFP. Maybe someday. For now, I know that this works well for my freshmen.
Really nice post Mark (as usual). You've given me a lot of food for thought on how I can make better use of this system in my own classes.
ReplyDeleteWe should talk about a teaching module, Paul. And maybe making some RFP and YFP based derivatives.
ReplyDelete